Words in brains
Alina Leinonen * & Christina M. Krause
Cognitive Science Unit, Department of Psychology, University of Helsinki
In this paper we review central methods in psycholinguistic research and discuss recent neurocognitive research in which language processing using Finnish has been studied. We further introduce different psycholinguistic research methods. Thereafter we discuss cortical correlates of language processing by introducing some main research results on event-related potentials. The use of neuroimaging methods with a high temporal resolution (such as EEG) with various experimental paradigms and analysis methods has proven useful in understanding the processing of language. Yet we still do not understand completely the nature of language related effects and what underlying neural mechanisms they manifest.
One of the intriguing questions in the study of human language processing is the way linguistic phenomena are represented and processed in the brain.
Since the 1950s, language processing has widely been studied in the field of psycholinguistics, mainly by means of behavioral experiments. In psycholinguistic experiments, subjects are typically presented with some form of linguistic input and asked to perform a task (e.g., make a judgment, name the stimulus, read a visually presented word aloud). The performance is usually measured using, e.g., reaction times (in milliseconds), eye-tracking, self-paced methods and/or error rates. Such behavioral measures give exact information about the speed of cognitive processing but give no information about how these processes are performed or where in the brain these processes are executed.
Psycholinguistic research generally involves core areas such as spoken word recognition, message construction, memory limitations, cross-linguistic comparisons and so on (for a review, see, e.g., MacWhinney 2001). Word recognition has been studied using, e.g., lexical decision tasks, various priming paradigms and naming tasks. Sentence comprehension has been studied using judgment tasks (e.g., correctness, semantic or grammatical judgments), such as judging syntactically or semantically anomalous and/or ambiguous sentences. Sentence production studies have involved, e.g., analysis of speech errors. For the past two decades brain imaging techniques have been widely used in association with psycholinguistic experimental tasks to get a better picture about language processing online (see below).
Today, many methods are available to study brain anatomy and, more interestingly, also brain activity simultaneously while the subject is performing cognitive, e.g., linguistic tasks. Both PET (positron emission tomography) and fMRI (functional magnetic resonance imaging) provide information about brain activity with a good spatial resolution. Thus, fMRI and PET show activation localized to specific regions, i.e., "where" activations take place in the brain. On the other hand, the temporal resolution of these methods is poor (between seconds and minutes). In contrast, the EEG (electroencephalogram) and the MEG (magnetoencephalogram) provide information about brain activity with a good temporal resolution, on the level of milliseconds. The spatial resolution of the EEG and MEG is, however, relatively poor. The EEG is one of the oldest psychophysiological methods (Berger 1929) but it is still successfully being used to assess brain activity, because it is a relatively inexpensive brain imaging method with an excellent temporal resolution.
In Finnish, the recognition and processing of complex words, e.g., inflected and derived words has been studied quite extensively using various behavioral methods such as visual lexical decision tasks and the eye-tracking method (e.g., Bertram et al. 1999, 2000, Laine et al. 1999, Niemi et al. 1994, Vannest et al. 2002). The mental processing of inflected words typically results in longer reaction times and higher error rates as compared to the processing of uninflected words. Thus, it has been proposed that most inflected Finnish words are processed via morpheme-based recognition, i.e., decomposed. It has been suggested that the decomposition process takes place in two stages (Niemi et al. 1994): First, the decomposition is performed at the visual word form level after which semantic and syntactic integration takes place.
Research on Finnish derivational morphology has suggested that derivational forms of the same stem are processed via their full forms, since participants perform equally well on monomorphemic and derived words in lexical decision tasks (e.g., Bertram et al. 1999, 2000, Niemi et al. 1994, Vannest et al. 2002, but see also Järvikivi et al. 2006). A recent reaction time study by Brattico et al. (2007) investigated the processing of polymorphemic novel Finnish words by means of varying the number of derivational suffixes. In this study (Brattico et al. 2007), novel Finnish words were generated by suffixing one to six derivational suffixes to the stems (e.g., "rosvo+sto+llinen" - a stem plus two derivational morphemes). The resulting stimulus words were completely novel to the subjects, as verified by a wide corpus search [1]. It was observed that reaction times increased linearly as the number of derivational suffixes increased, i.e., these novel words seemed to undergo morphological decomposition. When the number of derivational suffixes reached five, they were judged as nonwords, despite not violating any rules of Finnish word formation (for a more detailed description of the stimuli, see Brattico et al. 2007). Brattico et al. concluded that Finnish derivational process is most probably recursive but is regulated by factors such as working memory capacity.
On the basis of behavioral observations only, it has been difficult to test hypotheses concerning the neural representation and cortical processing of morphologically complex words. One reason for this is that it is difficult, if not impossible, to separate the influence of semantic, phonological, orthographic and morphological properties on, e.g., reaction time data (Morris & Holcomb 2005). Psychophysiological methods provide means to assess brain processes underlying various cognitive processes. Some of these methods - the EEG for instance - have a good temporal resolution which allow to study brain processes "online", simultaneously with, e.g., language processing. We will discuss the use of these methods and summarize some important research results below.
Event-related potentials provide a continuous, real-time measure of neural processing that is potentially sensitive to qualitatively different kinds of information (Kutas et al. 2000). Therefore, the ERPs have been used also by language researchers for the past two decades to uncover the time at which different types of information about words is processed. Event-related potentials are small voltage shifts in brain electric activity (EEG), which can be recorded from the scalp (see Figure 1 below). ERPs occur in association with motor, sensory and cognitive events and they are time locked to stimulus presentation. However, these modest electrical potentials are revealed only after averaging EEG samples over many stimulus repetitions. This summation operation removes the background activity not time-locked to the presentation of the stimulus, revealing the underlying response (Näätänen 1992). ERP components are typically identified by their polarity (positive or negative), latency (occurrence after the eliciting stimulus, in ms) and amplitude (in µV).
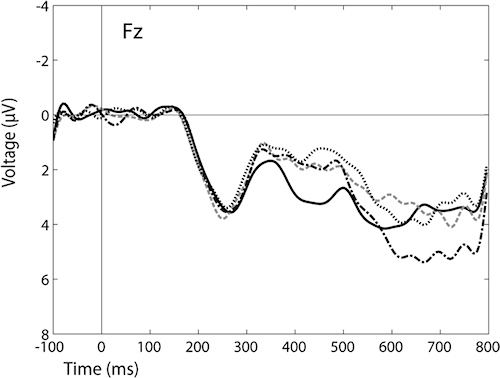
Figure 1. Grand average ERPs from electrode Fz, to critical words that were a) correct (solid line) words, b) inflectionally violated (dot line), c) derivationally violated (dash line) and d) combined violations (alternating dash/dot line), after baseline correction in the 100 ms prestimulus interval. Negative polarity is plotted upwards. X axis represents time (milliseconds), Y axis depicts voltage (microvolts, µV).
Several ERP components have been identified specifically in relation to language processing (for a review, see, e.g., Kutas et al. 2000). During visual and auditory word processing, it has been reported that phonetic processing has been reported at ~150-250 ms after stimulus onset (e.g., van Linden et al. 2007). Phonologically incongruent words have been reported to be observed at ~200-300 ms (Diaz & Swaab 2007, Van den Brink et al. 2001). These effects have been called as phonological mismatch negativity (PMN) (Diaz & Swaab 2007) and also the N200 effect (Van den Brink et al. 2001), which have been related to phonological processing.
Semantic deviations in sentences typically elicit a negative-going ERP component that peaks at around 400 ms after the onset of a semantically inappropriate word presented in a sentence, such as "I take coffee with cream and dog" (e.g., Kutas & Hillyard 1984, Kutas & Federmeier 2000, Kutas & Schmitt 2003). The N400 component has been interpreted to reflect relatively immediate online integration of a word's meaning into sentence semantic context (Hinojosa et al. 2001, Kutas & Schmitt 2003, Kotchoubey 2006). In addition to semantically violated sentences, the N400 effect has been robustly observed at word and discourse levels. Furthermore, besides semantic processing, the N400 amplitude has been observed to increase when processing a pronounceable pseudoword (Holcomb & Neville 1990), and thus the N400 effect has also been related to lexicality (whether a letter string is a word or not). In fact, it has been proposed that processing of almost any type of meaningful, or potentially meaningful, stimulus seems to be associated with the N400 effect.
Morphosyntactic violations (such as number and gender agreement violations) elicit the so-called left anterior negativity (LAN) ERP component, appearing at ~300-500 ms after the word onset and being most prominent at the frontal electrodes. The LAN has been reported also in association with syntactic agreement violations and word category constraints (Friederici 2002, Friederici & Weissenborn 2006). Thus, the LAN has been hypothesized to reflect a first stage of syntactic parsing (Friederici 2002). On the other hand, some authors have suggested that the LAN is generated in relation with gap-filling processing and thus has been interpreted as reflecting working-memory processing (King & Kutas 1995, Kluender & Kutas 1993). The functional interpretation of LAN effects is not yet agreed upon.
Various ERP studies on language have also reported a large positive waveform peaking at approximately 600 ms after the onset of words that are incongruous with the expected syntactic structure in garden path sentences, e.g., "The horse raced past the barn fell." (Osterhout & Holcomb 1992). This positive waveform has been called the P600 effect or Syntactic Positive Shift (SPS). It has been suggested that the amplitude of the P600 evoked by both syntactic ambiguities and violations could be modulated by both lexical and semantic contextual variables (Kuperberg 2007). The P600 effect has repeatedly been reported in association with processing of syntactically complex sentences (for recent reviews, see Friederici 2002, Friederici & Weissenborn 2006) and semantic/thematic violations in sentence context (Kuperberg 2007). The P600 effect has also been observed in association with inflectional (e.g., Lehtonen et al. 2007, Leinonen et al. 2008) and derivational (Leinonen et al. 2008) violations. A recent study using Finnish as study language reported of increased P600 amplitudes with increasing complexity of derived novel words (Brattico et al. submitted). The P600 effect has been interpreted as being related to repair of the violation and integrating it with the rest of the sentence (Friederici 2002).
It has been suggested that ERP observations on language processing show that ERP method is a power tool for tapping the time course of language comprehension (Kutas & Schmitt 2003). But based on the available ERP data, it seems plausible to assume that there is a significant amount of temporal overlap and interaction between various linguistic representations. Currently available data seems to be in agreement with the interactive models of word recognition: during word processing, different kinds of word-related information are activated in parallel, available after initial orthographic or phonemic analysis of a written or spoken word (e.g., Barber & Kutas 2007, Van den Brink & Hagoort 2004).
Language recognition and production involve complex information processing chains. Many researchers today agree on that memory, language, perception, attention, and other higher mental capacities are inseparable and the result of dynamic, distributed neural activity instead of the activity of strictly localized brain areas. Dynamic and distributed neural networks, oscillating at different frequencies are currently viewed as the most conceivable neural communication method. By means of such dynamic networks, large populations of neurons can interact, transform and process information and additionally constitute a unified picture of the external world in the mind. It has been noted that different frequencies in the EEG respond systematically differently to different cognitive processes and mental states and reflect alterations in, e.g., memory demands, language, attention, consciousness, stimulus modality, etc. (for recent reviews see, e.g., Knyazev 2007, Basar 2006, Ward 2003, Neuper and Pfurtscheller 2001, Basar et al. 2001b, Basar et al. 2000, Klimesch 1999, Pulvermüller et al. 1997, Basar et al. 1997, Krause 2006) (see Figure 2 for a depiction of EEG activity at different frequencies).
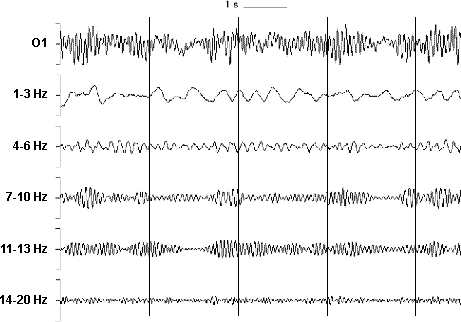
Figure 2. Examples of simultaneously occurring brain electric oscillations at 1-3 Hz (second row), 4-6 Hz (third row), 7-10 Hz (fourth row), 11-13 Hz (fifth row) and 14-20 Hz (sixth row) digitally filtered from the ongoing EEG (first row) from electrode O1. The Y axis depicts amplitude and the X axis stands for time. Note that the different frequencies are modulated differently as a function of time in relation to stimulus presentation points (vertical lines). Additionally, it can be seen that the amplitude of brain electric oscillations decreases with increasing frequency (adapted from Krause 2003).
Many recent studies indicate that language processing in the brain can also be studied by means of examining the functioning of brain oscillatory networks during linguistic processing (Bastiaansen et al. 2002a, 2002b, 2005, 2008, Hald et al. 2006; for a review, see Bastiaansen & Hagoort 2006). Language and memory are closely related and, for example, in semantic memory, which refers to the long-term memory of meanings, language plays a crucial role in shaping and maintaining memories. Therefore, language and memory can not necessarily be separated from each other.
Krause and co-workers have extensively investigated the encoding and recognition of acoustic verbal information utilizing the EEG (Krause et al. 1996, Krause et al. 1995, Krause et al. 2007, Krause et al. 2001). In these studies, it has repeatedly been shown that the encoding and recognition of auditorily presented linguistic material elicit distinct brain oscillatory responses. Thus, one can assess the neural circuits involved in verbal memory.
Brain oscillatory networks around 4 Hz seem to respond to word recognition and lexicality (Bastiaansen et al. 2005). Responses in the ~4 Hz frequency range have been reported in relation to processing of lexical-semantic information, e.g., pseudoword processing (Bastiaansen et al. 2008) as well as semantic and syntactic sentence comprehension (Bastiaansen et al. 2002a, 2002b, Hald et al. 2006). Leinonen et al. (2007) reported of delayed ~4 ERS responses during language switching when studying Finnish-Swedish bilinguals. It has been proposed that ~4-6 Hz responses are related to the retrieval of lexical-semantic information (Bastiaansen & Hagoort 2006), semantic retrieval and verbal working memory processing (Hald et al. 2006). Responses at ~4 Hz frequencies have also been related to the functioning of central executive attentional processes (Leinonen et al. 2007, Pesonen et al. 2006).
Brain oscillatory responses around ~10 Hz seem to respond to the retrieval of lexical-semantic information (Bastiaansen et al. 2008, Karrasch et al. 1998). For instance, processing of open and closed class words has elicited decrease in alpha power. Responses at ~10-12 Hz frequencies have been related to semantic memory processes (see Klimesch 1999 for a review). Thus, it may be the case that verbal content information is represented in memory by means of brain oscillatory networks operating specifically at ~10 Hz.
Responses around ~14-30 Hz frequency range have been observed as responses to syntactic filler-gap dependencies, which require a larger working memory load. It has been proposed that responses at ~14-30 Hz frequencies might indicate larger demands on verbal working memory and syntactic unification processes (Haarmann et al. 2002). It has been proposed that beta may be related to the efficiency of the processing of verbal material (Laine et al. under preparation). Some reports also exist on the effects of correct sentences (but not violated sentences) on brain oscillatory networks operating at higher frequencies, around 40 Hz (e.g., Hald et al. 2006). It has been suggested that (increases in) oscillatory activity in the gamma frequency range (~40 Hz) are part of the normal pattern of neural activity occurring during language processing (Hald et al. 2006).
The results from the studies on brain oscillations during linguistic processing are promising. It seems that it is possible to study the dynamics of the brain's language network by means of analyzing event-related changes in brain oscillations in a wide range of frequencies (Bastiaansen & Hagoort 2006). The ERPs and brain oscillations are two separate, but not necessarily independent properties of the brain. They are complementary to each other and offer a more comprehensive view of the dynamics of language processing in association with behavioral measures.
We do not yet know how information is perceived, coded, stored in and retrieved from the brain. Little is known about the neural basis underlying language processing. One of the reasons for this is that current experimental paradigms mostly violate some grammatical rules in order to elicit clear changes in the electrical activity of the brain. However, the violation types may not necessarily answer the questions that are raised by the researchers (Kutas et al. 2006). Moreover, we do not yet know the cognitive mechanisms that are manifested by the language-related ERP and oscillatory dynamic effects (Osterhout & Nicol 1999). Other limitations are related to the poor spatial or temporal resolution of brain imaging methods, which do not provide the whole picture about the processing of language in the brain. Recently, however, it begins to be possible to combine EEG and functional imaging methods and the future research seems promising.
It can be concluded that most probably language processing (as well as other cognitive processes) is a distributed property of the brain. It seems plausible to assume that language processing is executed by means of an interplay and interaction between sometimes distributed brain areas, by means of neuronal synchrony (Basar 2006, Basar et al. 1999). Thus, language and other higher cognitive processing might be enabled by means of an oscillating network, constantly evolving in time and space at different frequencies (Basar et al. 2000, Basar et al. 2001a, Cantero and Atienza 2005, Tallon-Baudry 2003, Pfurtscheller 2003, Neuper and Pfurtscheller 2001, Klimesch 1996).
Professor Christina M. Krause has been funded by the Academy of Finland (projects 7338, 42536, 1106071 and 1115215) and by the University of Helsinki (own research funds). Alina Leinonen is in the Biolinguistics graduate school funded by the University of Helsinki (own research funds).
* Corresponding author, address: Cognitive Science Unit, Department of Psychology, P.O. Box 9, FI-00014 University of Helsinki, Finland.
[1] A Finnish corpus composed by the Research Institute for the Languages of Finland, the Finnish IT Centre for Science and Department of General Linguistics, University of Helsinki. The corpus was used through WWW-Lemmie 2.0 at the Finnish IT Centre for Science, obtainable from . The corpus involved approximately 100 million word tokens.
Barber, H.A. & M. Kutas. 2007. "Interplay between computational models and cognitive electrophysiology in visual word recognition". Brain Research Reviews 53: 98-123. doi:10.1016/j.brainresrev.2006.07.002
Basar, E. 2006. "The theory of the whole-brain-work". International Journal of Psychophysiology 60: 133-138. doi:10.1016/j.ijpsycho.2005.12.007
Basar, E., C. Basar-Eroglu, S. Karakas & M. Schürmann. 1999. "Are cognitive processes manifested in event-related gamma, alpha, theta and delta oscillations in the EEG?". Neuroscience Letters 259: 165-168. doi:10.1016/S0304-3940(98)00934-3
Basar, E., C. Basar-Eroglu, S. Karakas & M. Schürmann. 2000. "Brain oscillations in perception and memory". International Journal of Psychophysiology 35: 95-124. doi:10.1016/S0167-8760(99)00047-1
Basar, E., C. Basar-Eroglu, S. Karakas & M. Schürmann. 2001a. "Gamma, alpha, delta and theta oscillations govern cognitive processes". International Journal of Psychophysiology 39: 241-248. doi:10.1016/S0167-8760(00)00145-8
Basar, E., M. Schürmann, C. Basar-Eroglu & S. Karakas. 1997. "Alpha oscillations in brain functioning: An integrative theory". International Journal of Psychophysiology 26: 5-29. doi:10.1016/S0167-8760(97)00753-8
Basar, E., M. Schürmann, T. Demiralp, C. Basar-Eroglu & A. Ademoglu. 2001b. "Event-related oscillations are 'real brain responses' - wavelet analysis and new strategies". International Journal of Psychophysiology 39: 91-127. doi:10.1016/S0167-8760(00)00135-5
Bastiaansen, M.C.M. & P. Hagoort. 2006. "Oscillatory neuronal dynamics during language comprehension". Progress in Brain Research 159: 179-196. doi:10.1016/S0079-6123(06)59012-0
Bastiaansen, M.C.M., R. Oostenveld, O. Jensen & P. Hagoort. 2008. "I see what you mean: Theta power increases are involved in the retrieval of lexical semantic information". Brain and Language 106: 15-28. doi:10.1016/j.bandl.2007.10.006
Bastiaansen, M.C.M., J.J.A. van Berkum & P. Hagoort. 2002a. "Syntactic processing modulates the theta rhythm of the human EEG". Neuroimage 17: 1479-1492. doi:10.1006/nimg.2002.1275
Bastiaansen, M.C.M., J.J.A. van Berkum & P. Hagoort. 2002b. "Event-related theta power increases in the human EEG during online sentence processing". Neuroscience Letters 323: 13-16. doi:10.1016/S0304-3940(01)02535-6
Bastiaansen, M.C.M., M. van der Linden, M. ter Keurs, T. Dijkstra & P. Hagoort. 2005. "Theta responses are involved in lexical-semantic retrieval during language processing". Journal of Cognitive Neuroscience 17: 530-541. doi:10.1162/0898929053279469
Berger, H. 1969 [1929]. "On the electroencephalogram of man". Hans Berger on the Electroencephalogram of Man: The Fourteen Original Reports on the Human Electroencephalogram, ed. by P. Gloor. Amsterdam: Elsevier.
Bertram, R., M. Laine, R.H. Baayen, R. Schreuder & J. Hyönä. 2000. "Affixal homonymy triggers full-form storage, even with inflected words, even in a morphologically rich language". Cognition 74: B13-B25. doi:10.1016/S0010-0277(99)00068-2
Bertram, R., M. Laine & K. Karvinen. 1999. "The interplay of word formation type, affixal homonymy, and productivity in lexical processing: Evidence from a morphologically rich language". Journal of Psycholinguistic Research 28: 213-226. doi:10.1023/A:1023200313787
Brattico, P., A. Leinonen, E. Brattico, M. Pekkola, V. Pehunen, A. Suni & C.M. Krause. Forthcoming. "The magnitude of the P600 component reflects lexical complexity". Submitted.
Brattico, P., A. Leinonen & C.M. Krause. 2007. "On the limits of productive word formation: Experimental data from Finnish". SKY Journal of Linguistics 20: 109-139. http://www.ling.helsinki.fi/sky/julkaisut/sky2007.shtml
Cantero, J.L. & M. Atienza. 2005. "The role of neural synchronization in the emergence of cognition across the wake-sleep cycle". Reviews in the Neurosciences 16: 69-83.
Diaz, M.T. & T.Y. Swaab. 2007. "Electrophysiological differentiation of phonological and semantic integration in word and sentence contexts". Brain Research 1146: 85-100. doi:10.1016/j.brainres.2006.07.034
Friederici, A.D. 2002. "Towards a neural basis of auditory sentence processing". Trends in Cognitive Sciences 6: 78-84. doi:10.1016/S1364-6613(00)01839-8
Friederici, A.D. & J. Weissenborn. 2006. "Mapping sentence form onto meaning: The syntax-semantic interface". Brain Research 1146: 50-58. doi:10.1016/j.brainres.2006.08.038
Haarmann, H.J., K.A. Cameron & D.S. Ruchkin. 2002. "Neural synchronization mediates on-line sentence processing: EEG coherence evidence from filler-gap constructions". Psychophysiology 39: 820-825. doi:10.1111/1469-8986.3960820
Hald, L.A., M.C.M. Bastiaansen & P. Hagoort. 2006. "EEG theta and gamma responses to semantic violations in online sentence processing". Brain and Language 96: 90-105. doi:10.1016/j.bandl.2005.06.007
Hinojosa, J.A., M. Martín-Loeches & F.J. Rubia. 2001. "Event-related potentials and semantics: An overview and an integrative proposal". Brain and Language 78: 128-139. doi:10.1006/brln.2001.2455
Holcomb, P.J. & H.J. Neville. 1990. "Auditory and visual semantic priming in lexical decision: A comparison using event-related brain potentials". Language and Cognitive Processes 5: 281-312. doi:10.1080/01690969008407065
Järvikivi, J., R. Bertram & J. Niemi. 2006. "Affixal salience and the processing of derivational morphology: The role of suffix allomorphy". Language and Cognitive Processes 21: 394-431. doi:10.1080/01690960400013213
Karrasch, M., C.M. Krause, M. Laine, A.H. Lang & M. Lehto. 1998. "Event-related desynchronization and synchronization during an auditory lexical matching task". Electroencephalography and Clinical Neurophysiology 107: 112-121. doi:10.1016/S0013-4694(98)00047-9
King, J.W. & M. Kutas. 1995. "The lexical processing negativity: An ERP whose latency indexes lexical characteristics of words". Psychophysiology 32: S45. doi:10.1111/j.1469-8986.1995.tb02379.x
Klimesch, W. 1996. "Memory processes, brain oscillations and EEG synchronization". International Journal of Psychophysiology 24: 61-100. doi:10.1016/S0167-8760(96)00057-8
Klimesch, W. 1999. "EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis". Brain Research Reviews 29: 169-195. doi:10.1016/S0165-0173(98)00056-3
Kluender, R. & M. Kutas. 1993. "Subjacency as a processing phenomenon". Language and Cognitive Processes 8: 573-633. doi:10.1080/01690969308407588
Knyazev, G.G. 2007. "Motivation, emotion, and their inhibitory control mirrored in brain oscillations". Neuroscience Biobehavioral Reviews 31: 377-395. doi:10.1016/j.neubiorev.2006.10.004
Kotchoubey, B. 2006. "Event-related potentials, cognition, and behavior: A biological approach". Neuroscience Biobehavioral Reviews 30: 42-65. doi:10.1016/j.neubiorev.2005.04.002
Krause, C.M. 2003. "Brain electric oscillations and cognitive processes". Experimental Methods in Neuropsychology, ed. by K. Hugdahl, 111-130. New York: Kluwer Academic Publishers.
Krause, C.M. 2006. "Cognition- and memory-related ERD/ERS responses in the auditory stimulus modality". Progress in Brain Research 159: 197-207. doi:10.1016/S0079-6123(06)59013-2
Krause, C.M., H. Lang, M. Laine, M. Kuusisto & B. Pörn. 1995. "Cortical processing of vowels and tones as measured by event-related desynchronization". Brain Topography 8: 47-56. doi:10.1007/BF01187669
Krause, C.M., H. Lang, M. Laine & B. Pörn. 1996. "Event-related EEG desynchronization and synchronization during an auditory memory task". Electroencephalography and Clinical Neurophysiology 98: 319-326. doi:10.1016/0013-4694(96)00283-0
Krause, C.M., M. Pesonen & H. Hämäläinen. 2007. "Brain oscillatory responses during the different stages of an auditory memory search task in children". Neuroreport 18: 213-216. http://journals.lww.com/neuroreport/pages/articleviewer.aspx?year=2007&issue=02120&article=00004&type=abstract
Krause, C.M., P-A. Salminen, I. Holopainen & L. Sillanmäki. 2001. "Event-related desynchronization and synchronization during a memory task in children". Clinical Neurophysiology 112: 2233-2240. doi:10.1016/S1388-2457(01)00684-8
Kuperberg, G. 2007. "Neural mechanisms of language comprehension: Challenges to syntax". Brain Research 1146: 23-49. doi:10.1016/j.brainres.2006.12.063
Kutas, M. & K.D. Federmeier. 2000. "Electrophysiology reveals semantic memory use in language comprehension". Trends in Cognitive Sciences 4: 463-470. doi:10.1016/S1364-6613(00)01560-6
Kutas, M., K.D. Federmeier, S. Coulson, J.W. King & T.F. Münte. 2000. "Language". Handbook of Psychophysiology, ed. by J.T. Cacioppo, L.G. Tassinary & G.G. Bentson, 576-601. Cambridge: Cambridge University Press. http://kutaslab.ucsd.edu/people/kutas/pdfs/2000.HP.pdf
Kutas, M. & S.A. Hillyard. 1984. "Brain potentials during reading reflect word expectancy and semantic association". Nature 307: 161-163. doi:10.1038/307161a0
Kutas, M. & B.M. Schmitt. 2003. "Language in microvolts". Mind, Brain, and Language: Multidisciplinary Perspectives, ed. by M.T. Banich & M. Mack, 171-209. Mahwah, N.J.: Lawrence Erlbaum Associates. http://kutaslab.ucsd.edu/people/kutas/pdfs/2003.MBL.171.pdf
Kutas, M., C. Van Petten & R. Kluender. 2006. "Psycholinguistics electrified II: 1994-2005". Handbook of Psycholinguistics, 2nd edition, ed. by M. Traxler & M.A. Gernsbacher, 659-724. New York: Elsevier. doi:10.1016/B978-012369374-7/50018-3
Laine, M., S. Hiltunen, R. Vigario, A-L. Säkkinen & C.M. Krause. Forthcoming. "Effects of controlled attention on simultaneous interpreters' working memory processes: EEG and reaction time study". Under preparation.
Laine, M., S. Vaino & J. Hyönä. 1999. "Lexical access routes to nouns in a morphologically rich language". Journal of Memory and Language 40: 109-135. doi:10.1006/jmla.1998.2615
Lehtonen, M., T. Cunillera, A. Rodríguez-Fornells, A. Hultén, J. Tuomainen & M. Laine. 2007. "Recognition of morphologically complex words in Finnish: Evidence from event-related potentials". Brain Research 1148: 123-137. doi:10.1016/j.brainres.2007.02.026
Leinonen, A., P. Brattico, M. Järvenpää & C.M. Krause. 2008. "Event-related potential (ERP) responses to violations of inflectional and derivational rules of Finnish". Brain Research 1218: 181-193. doi:10.1016/j.brainres.2008.04.049
Leinonen, A., M.O. Laine, M. Laine & C.M. Krause. 2007. "Electrophysiological correlates of memory processing in early Finnish-Swedish bilinguals". Neuroscience Letters 416: 22-27. doi:10.1016/j.neulet.2006.12.060
Linden, S. van, J.J. Stekelenburg, J. Tuomainen & J. Vroomen. 2007. "Lexical effects on auditory speech perception: An electrophysiological study". Neuroscience Letters 420: 49-52. doi:10.1016/j.neulet.2007.04.006
MacWhinney, B.J. 2001. "Psycholinguistics: Overview". International Encyclopedia of the Social & Behavioral Sciences, ed. by N.J. Smelser & P.B. Baltes, 12343-12349. Amsterdam: Elsevier. doi:10.1016/B0-08-043076-7/03024-2
Morris, J. & P.J. Holcomb. 2005. "Event-related potentials to violations of inflectional verb morphology in English". Cognitive Brain Research 25: 963-981. doi:10.1016/j.cogbrainres.2005.09.021
Näätänen, R. 1992. Attention and Brain Function. Hillsdale, N.J.: Lawrence Erlbaum Associates.
Neuper, C. & G. Pfurtscheller. 2001. "Evidence for distinct beta resonance frequencies in human EEG related to specific sensorimotor cortical areas". Clinical Neurophysiology 112: 2084-2097. doi:10.1016/S1388-2457(01)00661-7
Niemi, J., M. Laine & J. Tuominen. 1994. "Cognitive morphology in Finnish: Foundations of a new model". Language and Cognitive Processes 9: 423-446. doi:10.1080/01690969408402126
Osterhout, L. & P.J. Holcomb. 1992. "Event-related brain potentials elicited by syntactic anomaly". Journal of Memory and Language 31: 785-806. doi:10.1016/0749-596X(92)90039-Z
Osterhout, L. & J. Nicol. 1999. "On the distinctiveness, independence, and time course of the brain responses to syntactic and semantic anomalies". Language and Cognitive Processes 14: 283-317. doi:10.1080/016909699386310
Pesonen, M., C. Haarala, H. Hämäläinen & C.M. Krause. 2006. "Brain oscillatory 1-30 Hz EEG ERD/ERS responses during the different stages of an auditory memory search task". Neuroscience Letters 399: 45-50. doi:10.1016/j.neulet.2006.01.053
Pfurtscheller, G. 2003. "Induced oscillations in the alpha band: Functional meaning". Epilepsia 44(s12): 2-8. doi:10.1111/j.0013-9580.2003.12001.x
Pulvermüller, F., N. Birbaumer, W. Lutzenberger & B. Mohr. 1997. "High-frequency brain activity: Its possible role in attention, perception and language processing". Progress in Neurobiology 52: 427-445. doi:10.1016/S0301-0082(97)00023-3
Tallon-Baudry, C. 2003. "Oscillatory synchrony and human visual cognition". Journal of Physiology (Paris) 97: 355-363. doi:10.1016/j.jphysparis.2003.09.009
Van den Brink, D., C.M. Brown & P. Hagoort. 2001. "Electrophysiological evidence for early contextual influences during spoken-word recognition: N200 versus N400 effects". Journal of Cognitive Neuroscience 13: 967-985. doi:10.1162/089892901753165872
Van den Brink, D. & P. Hagoort. 2004. "The influence of semantic and syntactic context constraints on lexical selection and integration in spoken-word comprehension as revealed by ERPs". Journal of Cognitive Neuroscience 16: 1068-1084. doi:10.1162/0898929041502670
Vannest, J., R. Bertram, J. Järvikivi & J. Niemi. 2002. "Counterintuitive cross-linguistic differences: More morphological computation in English than in Finnish". Journal of Psycholinguistic Research 31: 83-106. doi:10.1023/A:1014934915952
Ward, L.M. 2003. "Synchronous neural oscillations and cognitive processes". Trends in Cognitive Sciences 7: 553-559. doi:10.1016/j.tics.2003.10.012
|
